Scalable, cost-efficient, and robust filtration and chromatography-based processes are required for the purification of adeno-associated virus (AAV) in bioprocessing. In this article, we show how we developed an efficient purification process for recombinant AAV (rAAV). The chromatography purification described includes affinity capture to maximize recovery with efficient impurity removal. The optimal elution conditions of the affinity step were different for rAAV2 and rAAV5 serotypes. The recovery for rAAV5 was significantly improved by removing the NaCl from the low pH elution buffer. We used anion exchange polishing to reduce the empty capsids and maximize enrichment of full rAAV capsids. In the polishing step, we developed an anion exchange protocol for rAAV5 where the empty capsids were first eluted with high MgCl2 concentration, and the full capsids eluted when an NaCl gradient was applied. The full capsid enrichment was 40% to 65% with a viral genome (VG) recovery of 60% to 80% depending on the pooling strategy used.
Introduction
The key factors for an effective AAV purification process are high overall yields and maximized empty capsid product impurity reduction. After harvest by detergent cell lysis and DNA degradation by nuclease treatment, the feed is clarified followed by a concentration and buffer exchange step that reduces the volume to prepare for the capture step. Affinity capture is an efficient way to specifically bind and concentrate the target and at the same time achieve a high impurity removal.
In this study, we adapted an affinity capture step for rAAV2 and rAAV5, followed by a polishing step with rAAV5 to improve the yield of rAAV full capsids (Fig 1). See how the upstream cell culture steps were carried out in these articles on upstream process development and rAAV production in suspension HEK293 cells with single-use bioreactors.
Capto™ AVB is an affinity chromatography resin designed for the purification of rAAV. The ligand of Capto™ AVB resin is a single domain antibody fragment that specifically binds rAAV 1, 2, 3, and 5 serotypes, and in addition genetically engineered recombinant variants depending on the presence of the AVB binding epitope region. The affinity ligand will not discriminate between the full rAAV capsid (containing the VG and gene of interest) and empty rAAV capsids (product-related impurity lacking viral genomes).
The proportion of full capsid product in the harvested material is usually well below 50%. High levels of empty capsids decrease efficacy and contaminate the product and thus need to be reduced. In addition to empty capsids, partially filled capsids containing truncated genomes or process-related impurities can be packed into the capsid during upstream production. Traditionally, CsCl gradient ultracentrifugation, which separates based on density differences, has been used to purify full rAAV capsids in small scale. However, for scalable manufacturing, chromatography-based methods are preferred.
Full capsids have a lower isoelectric point (pI) than empty capsids with approximate pI 5.9 vs 6.3 and the charge difference can be used to separate them by ion exchange chromatography using salt or pH elution. Moreover, other capsid properties of rAAV might affect the binding behavior depending on serotype. In cation exchange, using a lower pH buffer below capsid average pI, the capsid net charge is positive. Full capsids therefore elute first in a salt gradient since they are less positively charged compared to the empty capsids. If instead anion exchange is used, the capsid net charge is negative with a buffer pH higher than the average capsid pI. In this case, the empty capsids elute first in a salt gradient since they are less negatively charged compared to the full capsids (Fig 2). Additives such as detergents, carbohydrates, and metal ions can improve the separation.
To achieve accuracy of full and empty rAAV capsid analysis is challenging. However, it is critical to optimize the polishing step. Determining the percentage of full capsids using qPCR:ELISA relies on two independent assays, both with significant inherent variability, which results in uncertainty. We recommend using at least one additional orthogonal analytical method for percent full capsids to confirm results and increase accuracy.
In this study, we used the UV 260:280 ratio in the chromatograms to indicate the percentage of full capsids (approximately 0.6 to 0.7 for empty capsids and 1.1 to 1.3 for full capsids). To detect VG, we also determined the full and empty capsids ratio in the fractions by quantitative polymerase chain reaction (qPCR). We used ELISA to detect viral particles (VP) to determine full capsid percentage and VG recovery.
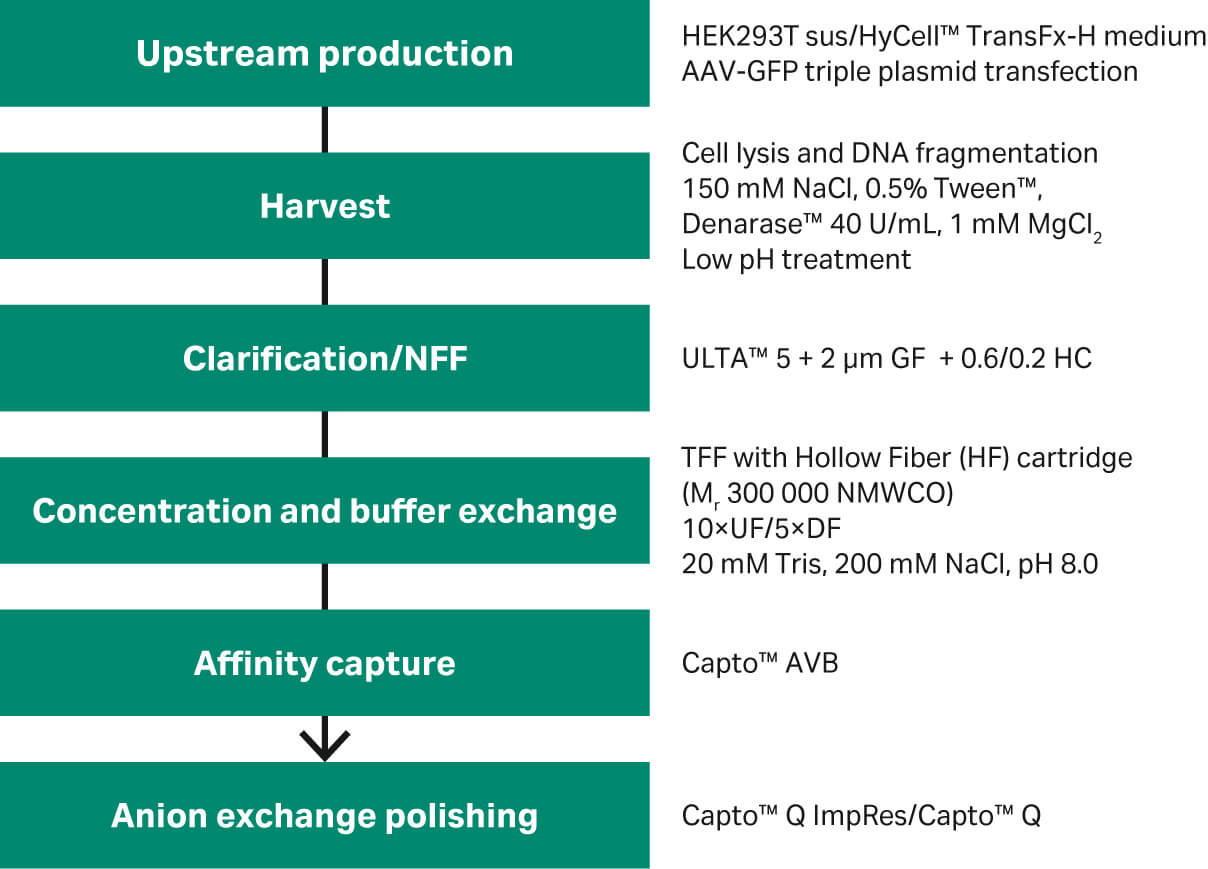
Fig 1. Overview of rAAV2 and rAAV5 purification steps.
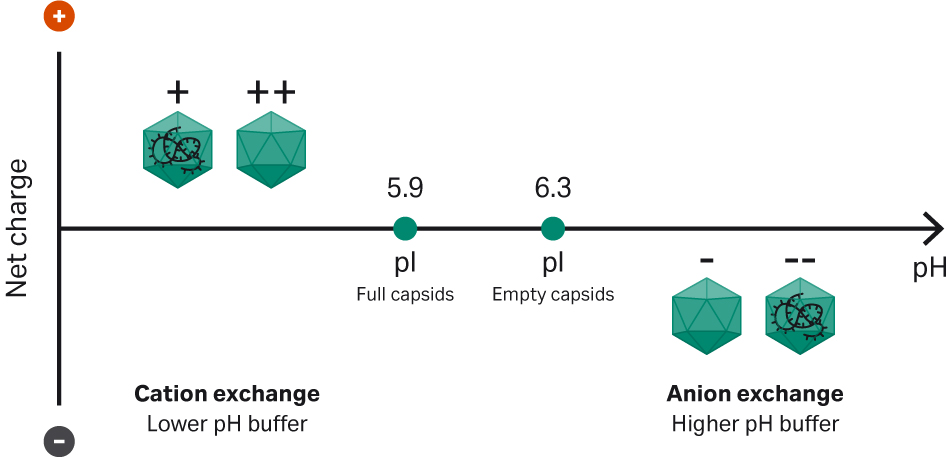
Fig 2. Principles of the separation of full and empty capsids using ion exchange.
Results and discussion
The starting materials for all experiments were obtained from rAAV production by triple plasmid transfection in HEK-293T cells, followed by harvest using detergent lysis (Tween™ 20), clarification, and concentration or buffer exchange by tangential flow filtration using hollow fibers (HF). For details, see Materials and Methods and Figure 1.
Optimizing the affinity capture step
The concentrated and buffer exchanged rAAV2, rAAV5, and rAAV8 samples were applied to affinity chromatography with Capto™ AVB and eluted in 50 mM citrate at pH 3.0 with gradient elution from 500 to 0 mM NaCl (Fig 3). rAAV2 is eluting completely at 500 mM NaCl. The rAAV5 eluted to some degree at high salt but the majority of the rAAV5 was eluted at the end of the gradient when the salt concentration was close to zero, which indicates that rAAV5 binds more strongly to the Capto™ AVB. It was clear that rAAV5 (but not rAAV2) recovery was improved as NaCl concentration in the buffer was reduced during elution at pH 3.0. The results in Figure 3 show that rAAV8 binds to a very low degree to Capto™ AVB.
Column: Capto™ AVB HiTrap™ column, 1 mL
Sample: rAAV2, 5, and 8 (TFF retentates)
Sample load: ~ 1 × 1013 VP/mL resin
Start buffer: 20 mM Tris buffer, 200 mM NaCl, pH 7.8
Elution buffer: 100 mM citrate pH 3.0, 500 to 0 mM NaCl gradient
Flow rate: 1 mL/min (150 cm/h)
System: ÄKTA™ pure 25
Detection: 280 nm
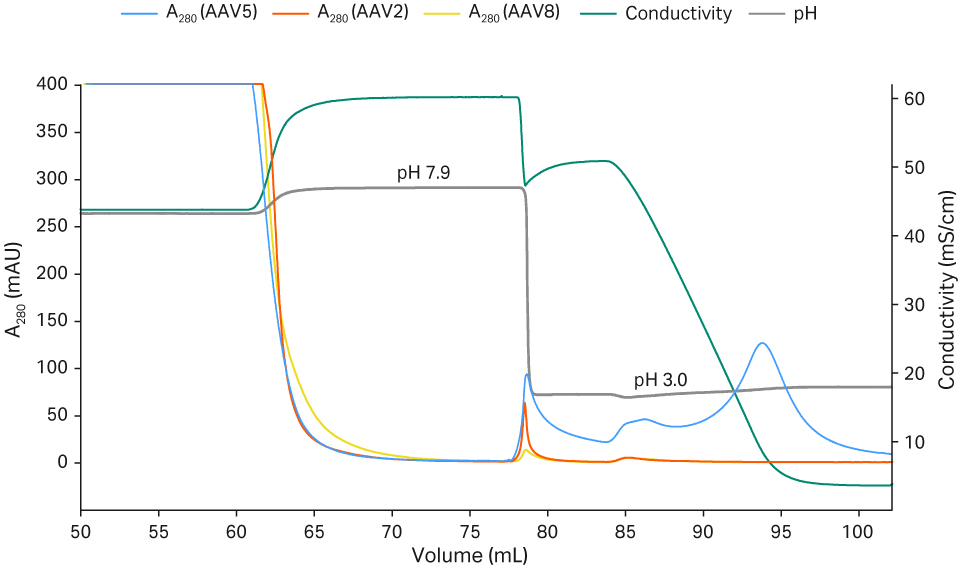
Fig 3. The effect of elution pH and elution strength for the recovery of rAAV2, 5, and 8.
A major part of the AAV5 did not elute using the AAV2 protocol (elution in 50 mM citrate pH 3.5, 500 mM NaCl, 500 mM arginine), since elution in the presence of NaCl reduced the recovery as seen in Figure 3. Thus, we continued to optimize the conditions for rAAV5 since this serotype was the focus for the process to be scaled up (see also scalable AAV production process).
We investigated if there were differences between 100 mM glycine and 100 mM citrate buffer with and without the addition of 500 mM NaCl (Fig 4). A concentration of 100 mM glycine without NaCl led to a more efficient elution at pH 3.0. This may be because of the higher buffer capacity of glycine at lower pH, but the conductivity is also increased which may reduce the elution of rAAV5 (Fig 3). So, to balance this, we chose these final elution conditions: 50 mM glycine and pH 2.7, which resulted in good VP recoveries (ranging from 60% to 90%). Elution with 500 mM NaCl regardless of the buffer used resulted in the poor recovery of rAAV5, even if the pH was reduced to 2.1 (Fig 4). The removal of arginine did not negatively affect the yields (analyzed with ELISA and transduction assay) of the eluted AAV5 (data not shown).
Column: Capto™ AVB HiTrap™ column, 1 mL
Sample: rAAV5 (TFF retentate)
Sample load: ~ 1 × 1013 VP/mL resin
Start buffer: 20 mM Tris buffer, 200 mM NaCl, pH 7.8 to 8.0
Elution buffer: 100 mM glycine or 100 mM citrate buffer with and without 500 mM NaCl
Flow rate: 1 mL/min (150 cm/h)
System: ÄKTA pure™ 25
Detection: 280 nm
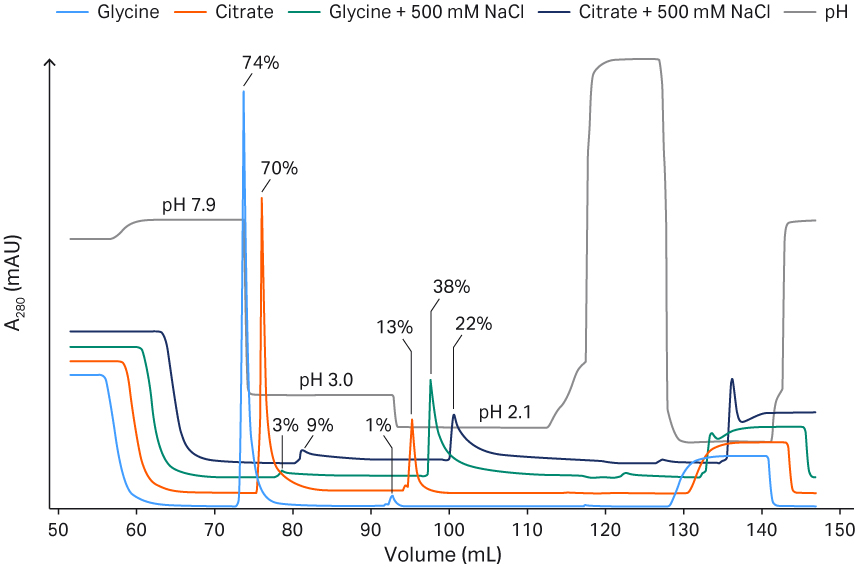
Fig 4. Comparison between glycine and citrate buffer and elution pH for Capto™ AVB with or without NaCl. The percentage recovery of rAAV5 total particles is indicated.
No full dynamic binding capacity (DBC) experiment was performed for rAAV2 but based on four high load experiments (ranging from 1.0 × 1014 to 1.8 × 1014 VP/mL of resin) with average VP recoveries of 83%. The highest load resulted in 100% VP recovery, so the DBC for rAAV2 was estimated to be above 2 × 1014 VP/mL resin.
Next, we needed to determine the DBC for rAAV5 using a classic breakthrough (BT) experiment approach, where a 1 mL Capto™ AVB HiTrap™ prepacked column was loaded until BT of rAAV5 occurred (Fig 5). The applied material contained 2.1 × 1012 VP/mL when analyzed with ELISA, and the 10% BT point corresponds to 3.8 × 1014 VP/mL of Capto™ AVB resin. The maximum capacity at 80% of the load to reach a 10% BT level was used. This corresponded to a DBC of 3.1 × 1014 rAAV5 particles/mL resin.
Column: Capto™ AVB HiTrap™ column, 1 mL
Sample: rAAV5
Sample load: 1.0 × 1014 to 1.8 × 1014 VP/mL of resin
Start buffer: 20 mM Tris buffer, 200 mM NaCl, pH 7.8
Equilibration buffer: 20 mM Tris buffer, 200 mM NaCl, pH 7.8
Flow rate: 1 mL (150 cm/h)
System: ÄKTA pure™ 25
Detection: 280 nm
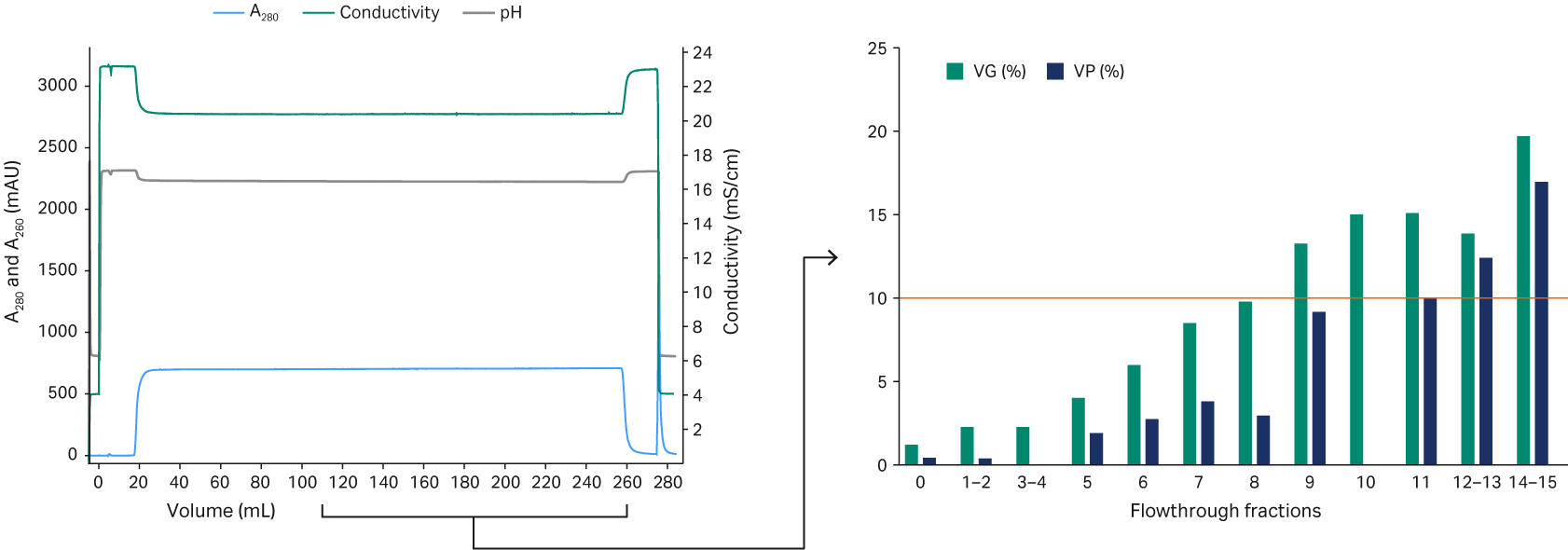
Fig 5. Breakthrough experiment using a 1 mL Capto™ AVB HiTrap™ chromatography column to determine DBC for rAAV5.
For rAAV2, a protocol (Fig 6) with elution in 50 mM citrate pH 3.5, 500 mM NaCl, 500 mM arginine was found optimal, resulting in high purity removal and average recovery of 90%. No further evaluations were performed for rAAV2.
A typical Capto™ AVB chromatogram (rAAV2) and the final detailed protocols can be seen in Figure 6. The high purity of the eluates for both rAAV2 and rAAV5 can be seen in Figure 7. The results are of a fluorescent multiplex Western blot detecting total protein or host cell protein (HCP) with Cy™5 prelabeling (similar sensitivity as silver staining) and viral proteins (VP1, VP2, and VP3) targeted by a primary antibody and Cy™3 labeled secondary antibody on the same blot. Cy5™ HCP signals could not be detected in the eluates, indicating good purity after Capto™ AVB chromatography.
No virus band was detected in the flowthrough fractions while clearly visible virus bands were detected in the eluate confirming ELISA results of approximately 100 to 200-fold concentration of rAAV in the affinity capture step. The transmission electron microscopy (TEM) image with the rAAV2 eluate also confirms high purity and single intact rAAV2 particles (Fig 7). The absence of salt in the elution buffer did not cause any aggregation of rAAV5 as evaluated from analytical size exclusion chromatography with Superose™ 6 Increase (< 1% data not shown).
Column: Capto™ AVB, HiTrap™ column, 1 mL
Sample: rAAV2 and 5
Sample load: ~ 170 to 215 CV (TFF retentate, 1 to 3 × 1014 VP/mL resin)
Start buffer: 20 mM Tris pH 7.8 + 200 mM NaCl, 10 CV
Elution Buffer: rAAV2: 50 mM citrate, pH 3.5, 500 mM NaCl, 500 mM arginine ,4 CV
rAAV5: 50 mM glycine pH 2.7, 5 CV
Flow rate: 1 mL/ min or 150 cm/h
System: ÄKTA pure™ 25
Detection: 260 and 280 nm
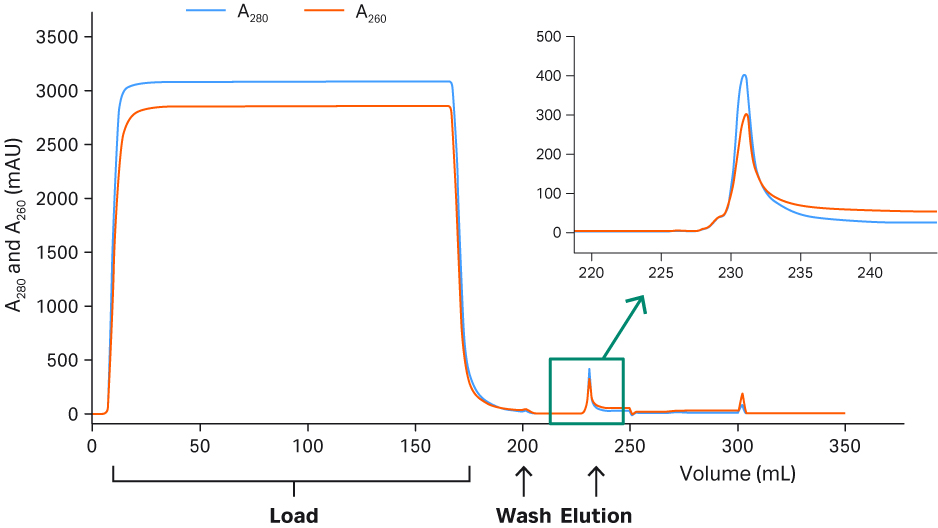
Fig 6. Chromatogram from the purification of rAAV2 on Capto™ AVB.
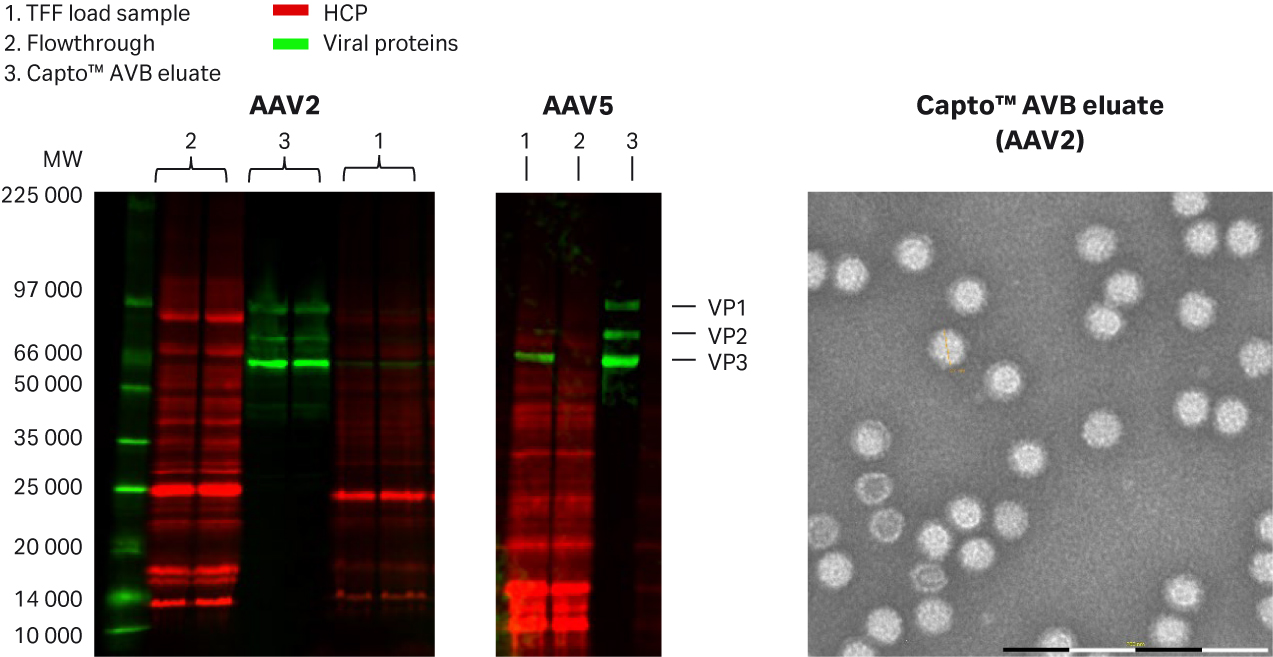
Fig 7. Purity analysis of Capto™AVB in (A) rAAV2, and (B) in rAAV5 eluates using multiplex fluorescence Western blot and (C) TEM image (MiniTEM™ from ViroNova) with negative staining. HCP was detected by Cy™5 prelabeling and rAAV proteins were targeted by primary anti AAV2 or AAV5 antibody and Cy™3 labeled secondary antibody.
Optimizing the polishing step
The purpose of the polishing step is to reduce the empty capsid product-related impurity as much as possible. The affinity-purified rAAV5 eluates were neutralized and diluted in equilibration buffer used in the experiment about ~ 10-fold to increase the pH and reduce the conductivity to between 1 and 3 mS/cm. We screened several resins for rAAV5 full and empty capsid separation performance (Table 1) and found that the anion exchange resins Capto™ Q ImpRes and Capto™ Q were the most promising candidates.
We selected Capto™ Q ImpRes for further optimization of running conditions and the results from the evaluation of are listed in Table 2.
Table 1. Screening of polishing resins for full and empty separation
| Resin | Sufficient rAAV5 binding | Sufficient VG/VP separation | VG recovery (%) |
| Capto™ Q ImpRes | Yes | Yes | 40 to 70 |
| Capto™ Q | Yes | Yes | 40 to 70 |
| Capto™ MMC ImpRes | Yes | No* | > 60 |
| Capto™ adhere ImpRes | Yes | No* | > 80 |
| Capto™ SP ImpRes | Yes | No* | > 80 |
*Using screening conditions
Table 2. Conditions and parameters tested with Capto™ Q ImpRes for full and empty separation
| Conditions and parameters tested | Comments |
| Gradient vs isocratic elution | Both possible, linear-gradient advantage for a robust recovery |
| Retention times (100 to 300 cm/h [2 and 6 min in HiScreen™ prepacked column]) | Similar results |
| pH 7.0 to 9.5 | High pH generally gave higher resolution |
| Buffer system (Tris or BTP) | Similar results, Tris is less expensive |
| MgCl2 (0 to 20 mM) | Critical parameter, 10 to 20 mM good range to evaluate |
| NaCl (up to 1 M) | Shallow gradient, small steps (elution below 0.5 M) |
| Additives (0.1% poloxamer 188 (nonionic surfactant), 1% sucrose) | Improved separation and analytical results |
Affinity purified rAAV5 sample was applied on Capto™ Q ImpRes equilibrated in 20 mM bis-Tris propane (BTP), pH 9.0. After a wash step, the same buffer containing 0, 5, 17, or 20 mM MgCl2 was applied followed by a shallow NaCl gradient (20 CV) up to 1 M NaCl containing the same constant concentration of MgCl2. All buffers contained additives (1% sucrose + 0.1% poloxamer 188).
A critical step was the use of an isocratic MgCl2 wash step to elute the empty capsids before an NaCl gradient to elute the full capsids. Increasing MgCl2 concentration causes elution of an increasing proportion of the empty capsids before the NaCl gradient (Fig 8 A to D). When the shallow NaCl gradient was applied, the full capsids eluted along with some empty capsids (Fig 8). A concentration of 20 mM MgCl2 is too high, since also some full capsids elute together with the empty capsids, resulting in reduced VG recovery (Fig 8D). The MgCl2 concentration needs to be fine-tuned to maximize the removal of empty capsids while retaining the full capsids for elution in the NaCl gradient. In this experiment, 17 mM MgCl2 (Fig 8C) improved separation compared to 20 mM MgCl2 (Fig 8D). A factor affecting the separation might be the ligand density of the anion exchange resin.
From the qPCR and ELISA results in Figure 8, we see that a high MgCl2 concentration can enhance the separation of full and empty particles and elute the full capsids earlier in the NaCl gradient.
The mechanism of how MgCl2 enhances the separation is unclear, but it may be due to the differential binding of Mg2+ ion between full and empty capsids, which in turn affects binding to the anion exchange ligand.
Column: Capto™ Q ImpRes HiScreen™ column, 4.7 mL
Sample: rAAV5 (Capto™ AVB eluate, 10-fold diluted in buffer A)
Sample load: 20 CV
Buffer A: 20 mM BTP or Tris pH 9.0 +/- additives (1% sucrose + 0.1% poloxamer 188)
Buffer B: 20 mM BTP or Tris pH 9.0 + 1 M NaCl + (0 to 20 mM) MgCl2 +/- additives (1% sucrose + 0.1% poloxamer 188) Flow velocity: 150 cm/h
Gradient: 1 mL/min
System: ÄKTA pure™ 25
Detection: 260 and 280 nm
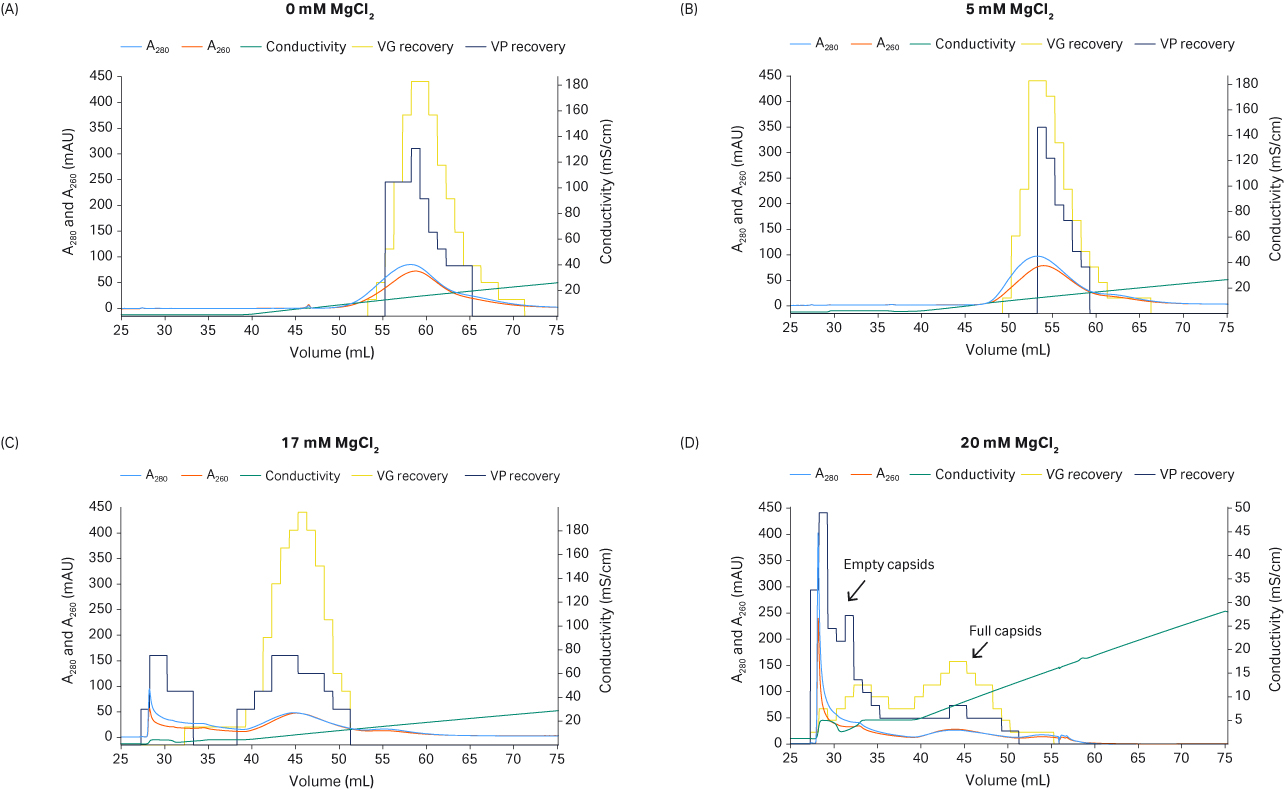
Fig 8. The effect of MgCl2 concentration on full and empty capsids separation. Recovery of VG (using qPCR) and VP (with ELISA) is shown in the yellow and dark blue histograms, respectively.
The separation of full and empty particles results in overlapping peaks, and there will be a trade off between the percentage of full capsids and VG recovery depending on how you pool the fractions. We used a procedure to help us define the pooling strategy (Fig 9). We were aiming for more than 50% full capsids and above 40% VG yield (Fig 9).
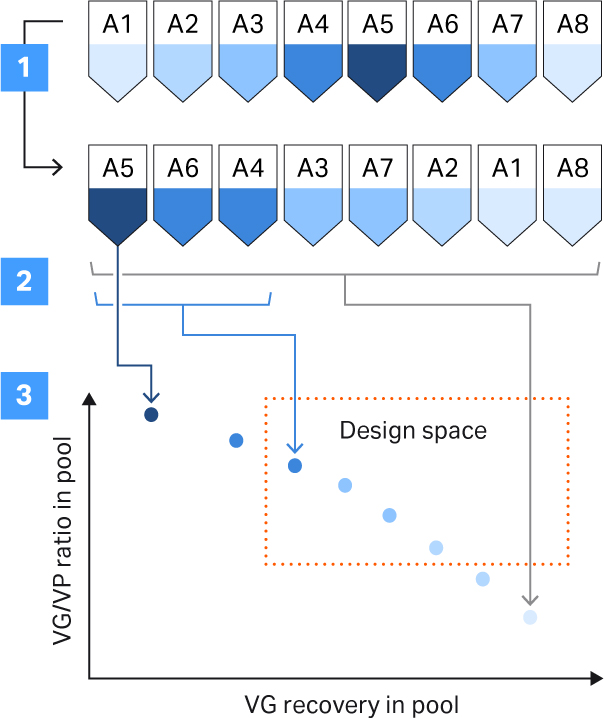
Fig 9. Procedure to support the pooling strategy. The steps involved were: (1) sorting of fractions based on full and empty capsid ratio; (2) calculation of full and empty capsid ratio and VG recovery in theoretical pools by including an increasing number of fractions in the pool; (3) plotting the calculated values for each pool and comparing them against the set criteria (design space, see the definitions section in this article to learn more about the design space).
The elution fractions were analyzed with qPCR and ELISA and sorted based on full and empty capsid ratios. Then, the full and empty ratio and VG recovery were calculated for the increasing number of fractions included in the pool. Our criteria for the design space were product and process criteria with above 50% full capsids, and 40% VG recovery defined by the final pooling strategy (Fig 9). The final protocol for separation of rAAV5 full and empty capsids using Capto™ Q ImpRes resulted in full capsid percentage between 40% and 65%. The VG recovery was between 60% and 80% depending on the pooling strategy (Fig 10). We determined capacity to be approximately 1 to 3 × 1013 VP/mL resin (data not shown).
Column: Capto™ Q ImpRes HiScreen™ column, 4.7 mL
Sample: rAAV5, Capto™ AVB HiTrap™, 1 mL eluate diluted 10-fold in Buffer A
Sample load: 5 CV (2 × 1012 VP/mL)
Buffer A: 20 mM Tris pH 9.0 + 18 mM MgCl2 + additives (1% sucrose and 0.1% poloxamer 188)
(Wash 1: Buffer A, 4 CV)
Buffer B: Buffer A + 1 M NaCl
Flow velocity: 150 cm/h or residence time 2.4 min
Gradient: Linear gradient (0% to 20% B), 15 CV. Elution hold at 0% Buffer B, 5 CV; elution hold at 20% Buffer B, 5 CV
System: ÄKTA pure™ 25
Detection: 260 and 280 nm
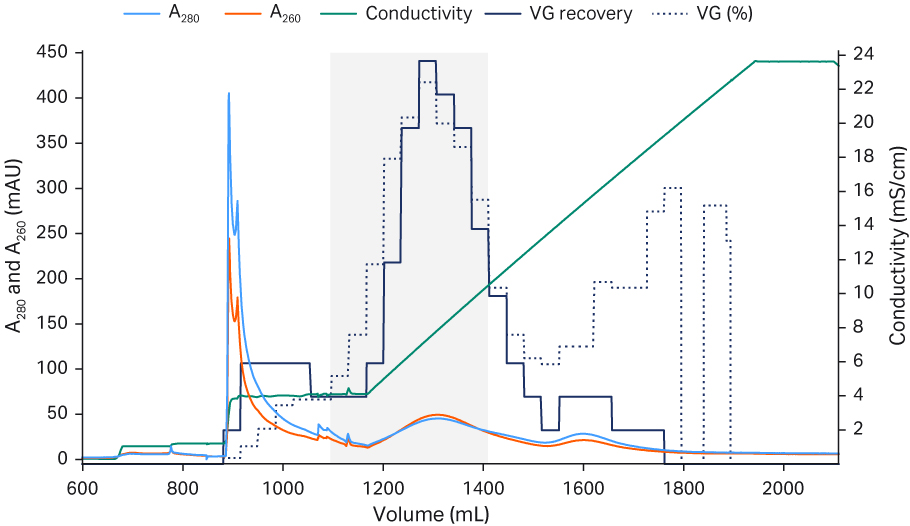
Fig 10. The final, optimized protocol for separation of rAAV5 full and empty capsids using Capto™ Q ImpRes resin
Gradient elution allows for small variations in conditions through the flexibility in pooling the correct fractions. However, an isocratic step elution protocol can simplify the process in large-scale GMP production. We have investigated step elution options for rAAV5 using Capto™ Q ImpRes (Fig 11) and Capto™ Q resins (Fig 12). Small isocratic step elutions can be optimized, varying the pH, MgCl2 and/or NaCl to achieve full- and empty-capsid separation. The results show a good separation between full and empty capsids. Capto™ Q resin contains dextran extenders, which can enhance separation also for other serotypes (work in progress).
Column: Capto™ Q ImpRes HiScreen™ column, 4.7 mL
Sample: rAAV5
Sample load: 5 CV
Buffer A: 20 mM Tris pH 9.0 + additives (1% sucrose and 0.1% poloxamer 188)
Buffer B: 20 mM Tris pH 9.0 + 10 mM MgCl2 and 1 M NaCl, pH 9.0
Flow velocity: 150 cm/h, residence time 2.4 min
Gradient: 11% to 100% Buffer B
System: AKTA pure™ 25
Detection: 280 and 260 nm
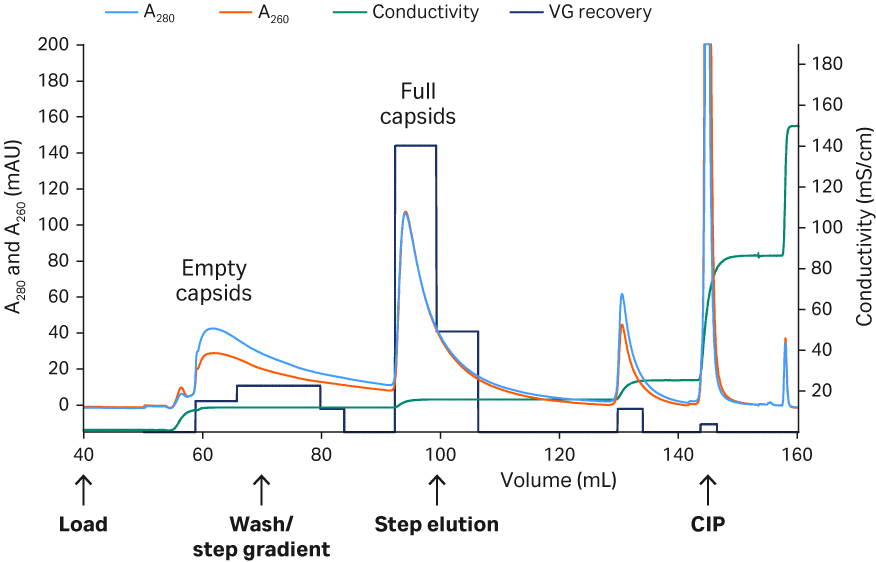
Fig 11. Separation of rAAV5 full and empty capsids with Capto™Q ImpRes resin and isocratic step elution (recovery of VG by qPCR is shown in dark blue histograms).
Column: Capto™ Q HiScreen™ column, 4.7 mL
Sample: rAAV5
Sample load: 5 CV
Buffer A: 20 mM BTP pH 9.5 + 2 mM MgCl2 + additives (1% sucrose + 0.1% poloxamer 188
Buffer B: 20 mM BTP pH 9.5 + 2 mM MgCl2 + additives (1% sucrose + 0.1% poloxamer188) + 1 M NaCl
Flow velocity: 150 cm/h, residence time 2.4 min
Gradient: 8% to 100% Buffer B
System: ÄKTA pure™ 25
Detection: 280 nm
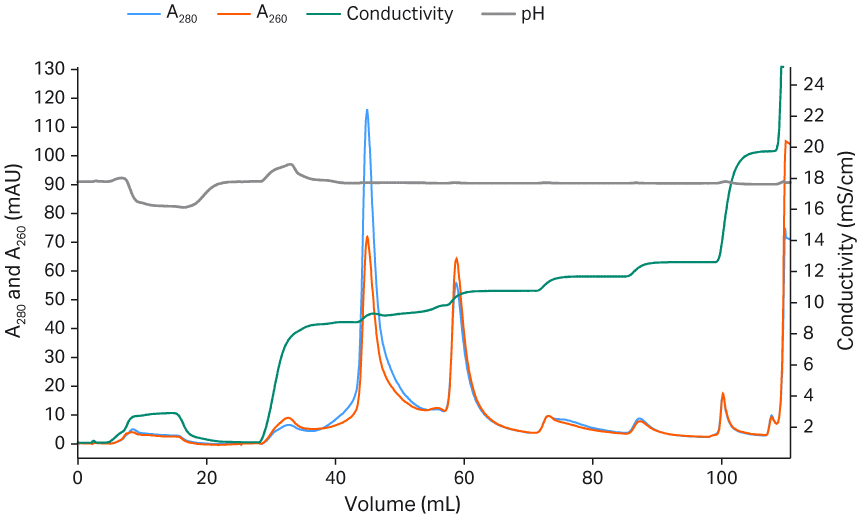
Fig 12. Separation of rAAV5 full and empty capsids with Capto™ Q resin and isocratic step elution.
Conclusions
- Capto™ AVB resin provided efficient affinity chromatography capture.
- Optimal Capto™ AVB elution conditions depended on the rAAV serotype used.
- Anion exchange polishing steps with Capto™ Q ImpRes or Capto™ Q resins can be used to enrich full capsids of AAV. MgCl2 concentration is critical in this step.
- Pooling strategy is key and a tradeoff between the ratio of full and empty capsids and yield of full capsids needs to be considered.
- The protocol can be transformed into isocratic step elution and is suitable for large-scale clinical grade production.
Acknowledgment
We thank iBET (Instituto de Biologia Experimental e Tecnológica), Oeiras, Portugal for affinity capture discussions and good collaboration.
CY24378-11Apr22-AN
TR29791259, 29792294, and 29646699
Materials and methods
Human embryonic kidney-293T CRL-3216 ATCC (HEK293T) cells adapted to suspension culture grown in HyClone™ HyCell™TransFx-H medium using Xcellerex™ XDR-10 and ReadyToProcess WAVE™ 25 bioreactors. An optimized triple plasmid (Rep/cap: helper: transgene GFP) transfection protocol was used to produce rAAV2-GFP or rAAV5-GFP. For further details see also AAV-associated epitopes and structure and dynamics of AAV serotypes.
Harvest and clarification
At the time of harvest, we lysed the cells with 0.5% Tween™ 20 + 150 mM NaCl and treated them with 40 U/mL Denarase™ nuclease (c-LEcta) + 1 mM MgCl2 for 4 h in the bioreactor. To avoid possible spontaneous precipitations, we treated the feed with pH 4.0 (by addition of 1M citric acid) for 30 min before neutralizing to pH 7.0 (by addition of 1M Tris base) and the resulting precipitate was allowed to sediment and discarded. The cell lysate was clarified by normal flow filtration (NFF) using a combination of 5 µm and 2 µm GF as well as 0.6 / 0.2 μm HC ULTA™ filters, each at approximately 500 cm2 /L harvest feed ( see rAAV production).
Tangential flow filtration (TFF)
The clarified feed was concentrated 10-fold by TFF on a ReadyToProcess™ hollow-fiber filter (UFP-300-C-4x2MA) using an ÄKTATM flux 6 tangential flow filtration system. The filter area of this filter cartridge is 1400 cm2 with a NMWC of Mr 300,000. The membrane flow rate was approximately 140 cm2/L clarified feed with a shear rate of 7 to 9000 s-1 and a transmembrane pressure (TMP) of 0.09 MPa (0.9 bar, 13.1 psi). After concentration, a five-fold volume buffer exchange was performed to 20 mM Tris pH 7.8, 200 mM NaCl ( see also rAAV production).
Affinity capture
We used Capto™ AVB resin prepacked in 1 or 5 mL HiTrap™ columns for affinity capture. The columns were connected to ÄKTA™ pure 25 chromatography system for the capture of recombinant rAAV2 and rAAV5. The residence time was about 150 cm/h for all the steps. We used either glycine or citrate buffer, and a detailed protocol can be seen in Figure 6. The load was approximately 1 to 3 × 1014 VP/mL resin.
Polishing
Capto™AVB resin eluate was diluted approximately 10-fold in equilibration buffer (composition depending on the experiment). We used Capto™ Q ImpRes resin packed in a 2 mL Tricorn™ 5/100 and Capto Q resin prepacked in 4.7 mL HiScreen™ column connected to ÄKTA™ pure 25 chromatography system for all the experiments apart from the large-scale run with the final optimized protocol where a 51 mL HiScale™ 26 column was used. The load was approximately 1 × 1013 VP/mL resin.
Tris or Bis-tris propane (BTP) buffers were used. The residence time was 2.4 min with a flow velocity of 250 cm/h for all steps.
Analytics
Total rAAV particle VP titer was determined using AAV2 or AAV5 ELISA (Progen). VG titer was analyzed using qPCR targeting the CMV (Cytomegalovirus) sequence. Multiplex fluorescence was performed using Cy™5 prelabeling reagent (Cytiva), primary anti-AAV antibody (Progen), and Cy™3-labeled secondary antibody (Cytiva). SDS-PAGE and Western blot analysis were used to analyze fractions from Capto™ AVB. TEM of rAAV2 was performed with negative staining using a MiniTEM™ system, Vironova AB (see results in Fig 7).